Liming Liang,
Wenxiang Bi,
Yuanyuan Tian ![]()
For correspondence:- Yuanyuan Tian Email: ianyuan@sdu.edu.cn Tel:+8653188382092
Received: 9 October 2015 Accepted: 11 March 2016 Published: 30 April 2016
Citation: Liang L, Bi W, Tian Y. Autophagy in photodynamic therapy. Trop J Pharm Res 2016; 15(4):885-889 doi: 10.4314/tjpr.v15i4.31
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Macroautophagy (autophagy) is crucial for cell survival during starvation and plays important roles in human diseases. It is a highly conserved intracellular degradation system in eukaryotes for removal and recycling of cytoplasmic components including damaged proteins and organelles to obtain energy. The relationship between cancer and autophagy has been extensively studied in recent years. In cancer and cancer therapy, autophagy acts as a double-edged sword. Photodynamic therapy (PDT) is a kind of tumor therapy applied with a tumor-localizing photosensitizing agent which is followed by activation with the light of a specific wavelength. How much is autophagy involved in photodynamic therapy? The work in this area is still limited.
Introduction
Autophagy is a conserved intracellular degradation process in which cellular organelles, proteins and invading microbes are degraded by lysosomes. There are three types of autophagy: macroautophagy, mitoautophagy and chaperone-mediated autophagy. This review is focused on macroautophagy which is referred to as autophagy hereafter. It is induced by inactivation of mTOR complex 1 (mTORC1) and regulated by a series of Atg proteins. It performs double roles: promotes cell survival or cell death, also in cancer therapies [1].
Activation of autophagy is found in some human primary tumors. Autophagy protects cells against shortage of nutrients. The amino acids and fatty acids produced by autophagic degradation are used to generate ATP to help cancer cells to survive in an unfavorable starved environment [2].
APOPTOSIS AND AUTOPHAGY
Apoptosis is an energy dependent cell-programmed death process in nucleated cells. Apoptosis, different from cell necrosis that is accompanied with severe irreversible injury, means a selective removal of individuals without complete disruption of the tissue. Apoptosis is recognized as an important cellular event during both normal development and disease progression [3,4]. During apoptosis, the intracellular content is wrapped in a small bubble of the membrane and swallowed by phagocyte and it is harmless to the surrounding cells. Apoptosis possesses unique morphologic features such as intact karyotheca, condensed nuclear chromatin attached to the karyotheca, wide perinuclear spaces, swollen mitochondria, formation of apoptotic bodies and cell shrinkage [5,6].
Caspases are responsible for many of the biochemical and morphological changes associated with apoptosis. Pathways for induction of apoptosis have been identified—intrinsic and extrinsic, one involving caspase-8 and the other involving Caspase-9 as the most apical caspase, which can activate effector caspase, such as Caspase-3, -6, -7. The effector caspases then cleave intracellular substrates, thereby important cellular processes are disabled and eventually cell death is caused [7-9].
Autophagy is another kind of programmed cell death in addition to apoptosis. It is a self-degradative catabolic process by which cells digest themselves. Autophagy is a fundamental function of eukaryotic cells and is well conserved from yeast to humans. The most typical trigger of autophagy is nutrient starvation; in this sense, lack of any type of essential nutrient can induce autophagy. In yeast, nitrogen starvation is the most potent stimulus [10].
Autophagosomes are double-membrane cytoplasmic vesicles that can engulf various cellular constituents, and then autophagosomes fuse with lysosomes to form autolysosomes, where sequestered cellular components are digested [11].
The autophagosomes are usually ultrastructures visualized by transmission electron microscope (TEM), and so it is a gold-standard method for determination of autophagy [12]. Although TEM has been the gold standard, this method requires considerable skills. Recent studies of autophagy have been expanded to the marker proteins of autophagy, and the molecular mechanism is now well understood.
mTOR, Atg proteins and LC3
Recent research has revealed roles of the protein kinase termed ‘the target of rapamycin' (TOR) in autophagy. The mammalian target of rapamycin, mTOR, interacts with other proteins to form two main types of complex, mTOR complexes 1 and 2 (mTORC1 and mTORC2) [13], which can regulate autophagy negatively. In yeast, 31 autophagy-related (Atg) proteins have been identified, and many of them gather at a site that can be identified by fluorescence microscopy [10,14]. In mammalian, Beclin-1 and light chain 3 (LC3) are homologous proteins of Atg6 and Atg8 protein. Classical autophagy initiation begins with the complex involving Beclin-1. It is now widely known that microtubule-associated protein LC3 is related to autophagy monitoring. The conversion of LC3-I to LC3-II is indicative of autophagic activity.
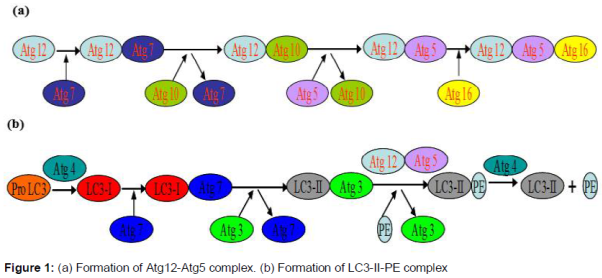
Atg12-Atg5 complex
Atg12 is conjugated to Atg5, catalyzed by two enzymes, Atg7 and Atg10. In this process, Atg proteins are modified and catalyzed in a ubiqutin-like way. Then Atg12-Atg5 conjugate associate with Atg16 and this contributes to bring LC3-II and phosphatidylethanolamine (PE) together to form LC3-II-PE complex () [15].
Formation of LC3-II
Pro-LC3 is turned into LC3-I catalyzed by Atg4 and Atg7, and then LC3-II-PE complex is formed catalyzed by Atg3, Atg12-Atg5 complex, finally free LC3-II is released () [16]. Soluble LC3-I and lipid bound LC3-II are associated with the formation of autophagosomes.
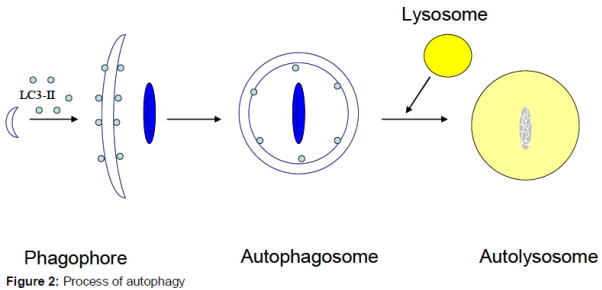
Process of autophagy
In the initial stage, the autophagosome, a double-membrane vesicle, arises from a membrane structure named phagophore which extends and sequesters cytoplasmic organelles such as mitochondria, endoplasmic reticulum and ribosomes, and then the edges of the membrane fuse to form the spherical structure, autophagosome. After fusing with lysosomes, the inner membrane and the engulfed components of the autophagosome are degraded by lysosome, forming a single-membrane vacuole structure called autolysosome () [17,18].
Cell death in photodynamic therapy
Photodynamic therapy can induce apoptosis, necrosis and autophagy, which is affected by many factors, such as the types of photosensitizers, cells and photodynamic dosage. The porphyrin family molecules, such as 5-ALA which localize in the mitochondria induce generation of reactive oxygen species (ROS) [19,20] and mainly result in mitochondrial apoptosis involving activation of caspase cascades after photoactivation [21]. Autophagy is detected following PDT mediated by photosensitizers targeted at endoplasmic reticulum [22-24], mitochondria [25,26] or both endoplasmic reticulum and mitochondria [27].
Sasnauskiene’s studies of Safr-mediated PDT on A-431 cells revealed that damage to mitochondrial interior at low and intermediate photodynamic dosages did not result in apoptosis; it induced autophagy in cells. However, when treated with a higher photodynamic dosage, extensive apoptosis appeared besides autophagy [28]. Kessel et al recently reported that L1210 cells underwent both autophagy and apoptosis following PDT with the endoplasmic reticulum sensitizer CPO, similar processes occurred in L1210 cultures following PDT with the mitochondrial sensitizer MC [29]. Recent observation of Francois et al demonstrated that cells submitted to the lowest PDT-dose displayed significant LC3-II expression, but there was no pronounced cyt C release and caspase cleavage [30].
Due to the high reactivity of photogenerated ROS [31-33], autophagy is initiated to remove oxidatively damaged organelles, such as mitochondria and endoplasmic reticulum which are targets of the photosensitizers [21,34,35]. Previous findings reflect that autophagy contributes to cell survival; in addition, antitumorigenic roles of autophagy are also mentioned in some research. A combination of autophagy inhibitor with PDT could promote apoptotic death, thus enhancing the treatment effect [36]. Therefore, autophagy can protect cells and help them to tolerate photodynamic therapy; however, if there is a high level of autophagy, it will lead to cell death.
Declarations
Acknowledgement
References
Archives


News Updates